Retinal (also known as retinaldehyde) is a polyene chromophore. Retinal, bound to proteins called opsins, is the chemical basis of visual phototransduction, the light-detection stage of visual perception (vision).


Some microorganisms use retinal to convert light into metabolic energy.
There are many forms of vitamin A — all of which are converted to retinal, which cannot be made without them. Retinal itself is considered a form of vitamin A when eaten by an animal. The number of different molecules that can be converted to retinal varies from species to species. Retinal was originally called retinene,[2] and renamed[3] after it was discovered to be vitamin A aldehyde.[4][5]
Vertebrate animals ingest retinal directly from meat, or they produce retinal from carotenoids — either from α-carotene or β-carotene — both of which are carotenes. They also produce it from β-cryptoxanthin, a type of xanthophyll. These carotenoids must be obtained from plants or other photosynthetic organisms. No other carotenoids can be converted by animals to retinal. Some carnivores cannot convert any carotenoids at all. The other main forms of vitamin A — retinol and a partially active form, retinoic acid — may both be produced from retinal.
Invertebrates such as insects and squid use hydroxylated forms of retinal in their visual systems, which derive from conversion from other xanthophylls.
Vitamin A metabolism
Living organisms produce retinal (RAL) by irreversible oxidative cleavage of carotenoids.[6]
For example:
- beta-carotene + O2 → 2 retinal,
catalyzed by a beta-carotene 15,15'-monooxygenase[7] or a beta-carotene 15,15'-dioxygenase.[8]
Just as carotenoids are the precursors of retinal, retinal is the precursor of the other forms of vitamin A. Retinal is interconvertible with retinol (ROL), the transport and storage form of vitamin A:
- retinal + NADPH + H+ ⇌ retinol + NADP+
- retinol + NAD+ ⇌ retinal + NADH + H+,
catalyzed by retinol dehydrogenases (RDHs)[9] and alcohol dehydrogenases (ADHs).[10]
Retinol is called vitamin A alcohol or, more often, simply vitamin A. Retinal can also be oxidized to retinoic acid (RA):
- retinal + NAD+ + H2O → retinoic acid + NADH + H+ (catalyzed by RALDH)
- retinal + O2 + H2O → retinoic acid + H2O2 (catalyzed by retinal oxidase),
catalyzed by retinal dehydrogenases[11] also known as retinaldehyde dehydrogenases (RALDHs)[10] as well as retinal oxidases.[12]
Retinoic acid, sometimes called vitamin A acid, is an important signaling molecule and hormone in vertebrate animals.
Vision
Retinal is a conjugated chromophore. In the human eye, retinal begins in an 11-cis-retinal configuration, which — upon capturing a photon of the correct wavelength — straightens out into an all-trans-retinal configuration. This configuration change pushes against an opsin protein in the retina, which triggers a chemical signaling cascade, which can result in perception of light or images by the human brain. The absorbance spectrum of the chromophore depends on its interactions with the opsin protein to which it is bound, so that different retinal-opsin complexes will absorb photons of different wavelengths (i.e., different colors of light).
Opsins

Opsins are proteins and the retinal-binding visual pigments found in the photoreceptor cells in the retinas of eyes. An opsin is arranged into a bundle of seven transmembrane alpha-helices connected by six loops. In rod cells, the opsin molecules are embedded in the membranes of the disks, which are entirely inside of the cell. The N-terminus head of the molecule extends into the interior of the disk, and the C-terminus tail extends into the cytoplasm of the cell. In cone cells, the disks are defined by the cell's plasma membrane, so that the N-terminus head extends outside of the cell. Retinal binds covalently to a lysine on the transmembrane helix nearest the C-terminus of the protein through a Schiff base linkage. Formation of the Schiff base linkage involves removing the oxygen atom from retinal and two hydrogen atoms from the free amino group of lysine, giving H2O. Retinylidene is the divalent group formed by removing the oxygen atom from retinal, and so opsins have been called retinylidene proteins.
Opsins are prototypical G protein-coupled receptors (GPCRs).[13] Bovine rhodopsin, the opsin of the rod cells of cattle, was the first GPCR to have its X-ray structure determined.[14] Bovine rhodopsin contains 348 amino acid residues. The retinal chromophore binds at Lys296.
Although mammals use retinal exclusively as the opsin chromophore, other groups of animals additionally use four chromophores closely related to retinal: 3,4-didehydroretinal (vitamin A2), (3R)-3-hydroxyretinal, (3S)-3-hydroxyretinal (both vitamin A3), and (4R)-4-hydroxyretinal (vitamin A4). Many fish and amphibians use 3,4-didehydroretinal, also called dehydroretinal. With the exception of the dipteran suborder Cyclorrhapha (the so-called higher flies), all insects examined use the (R)-enantiomer of 3-hydroxyretinal. The (R)-enantiomer is to be expected if 3-hydroxyretinal is produced directly from xanthophyll carotenoids. Cyclorrhaphans, including Drosophila, use (3S)-3-hydroxyretinal.[15][16] Firefly squid have been found to use (4R)-4-hydroxyretinal.
Visual cycle
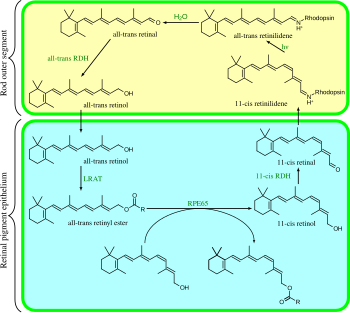
The visual cycle is a circular enzymatic pathway, which is the front-end of phototransduction. It regenerates 11-cis-retinal. For example, the visual cycle of mammalian rod cells is as follows:
- all-trans-retinyl ester + H2O → 11-cis-retinol + fatty acid; RPE65 isomerohydrolases;[17]
- 11-cis-retinol + NAD+ → 11-cis-retinal + NADH + H+; 11-cis-retinol dehydrogenases;
- 11-cis-retinal + aporhodopsin → rhodopsin + H2O; forms Schiff base linkage to lysine, -CH=N+H-;
- rhodopsin + hν → metarhodopsin II (i.e., 11-cis photoisomerizes to all-trans):
- (rhodopsin + hν → photorhodopsin → bathorhodopsin → lumirhodopsin → metarhodopsin I → metarhodopsin II);
- metarhodopsin II + H2O → aporhodopsin + all-trans-retinal;
- all-trans-retinal + NADPH + H+ → all-trans-retinol + NADP+; all-trans-retinol dehydrogenases;
- all-trans-retinol + fatty acid → all-trans-retinyl ester + H2O; lecithin retinol acyltransferases (LRATs).[18]
Steps 3, 4, 5, and 6 occur in rod cell outer segments; Steps 1, 2, and 7 occur in retinal pigment epithelium (RPE) cells.
RPE65 isomerohydrolases are homologous with beta-carotene monooxygenases;[6] the homologous ninaB enzyme in Drosophila has both retinal-forming carotenoid-oxygenase activity and all-trans to 11-cis isomerase activity.[19]
Microbial rhodopsins
All-trans-retinal is also an essential component of microbial, opsins such as bacteriorhodopsin, channelrhodopsin, and halorhodopsin. In these molecules, light causes the all-trans-retinal to become 13-cis retinal, which then cycles back to all-trans-retinal in the dark state. These proteins are not evolutionarily related to animal opsins and are not GPCRs; the fact that they both use retinal is a result of convergent evolution.[20]
History
The American biochemist George Wald and others had outlined the visual cycle by 1958. For his work, Wald won a share of the 1967 Nobel Prize in Physiology or Medicine with Haldan Keffer Hartline and Ragnar Granit.
| This article uses material from the Wikipedia article Metasyntactic variable, which is released under the Creative Commons Attribution-ShareAlike 3.0 Unported License. |